![]()
Aroids and other genera in the Collection
Take the Tour Now?
Orchids
The
Exotic Rainforest
Images on this website are copyright protected. Contact
us before attempting to reuse.
New:
Understanding, pronouncing and using
Botanical terminology, a Glossary
Lois is an aroid and this page explains what you are seeing.
http://www.hmnsmedia.org/CorpseFlower/
Photo of Lois below
including
Anthurium, Philodendron, Alocasia,
Amorphophallus, Colocasia, Spathiphyllum, Monstera
and other aroid genera
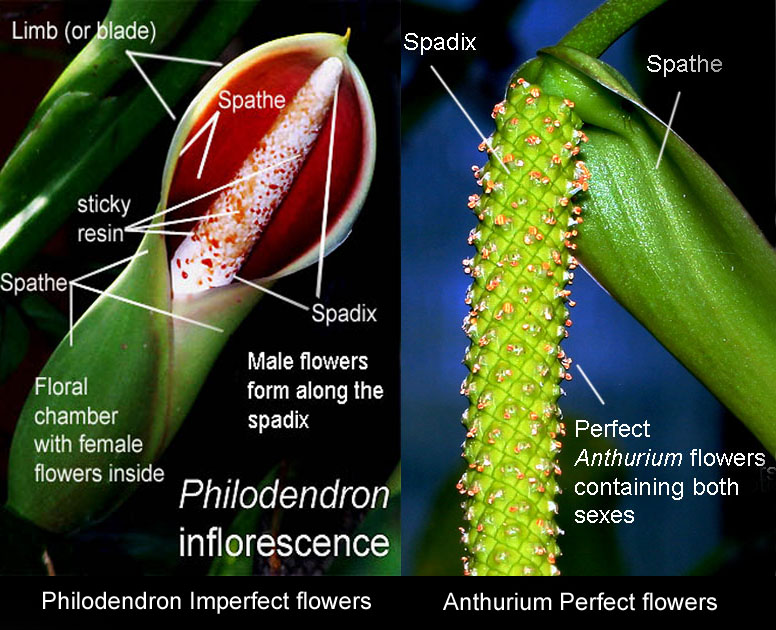
By Julius Boos
with additional input by Steve Lucas

 The
spathe
appears in the
shape of a hood while the spadix
that holds the flowers is
a spike on a thickened fleshy axis.
In
botany a bract is a specialized leaf that is
associated with the reproductive structure
of the plant. The spadix can only
produce flowers at sexual anthesis.
True flowers contain
near microscopic sexual parts including
anthers, stamens, and stigmas. Some
aroids have tepals but others do not.
The
spathe
appears in the
shape of a hood while the spadix
that holds the flowers is
a spike on a thickened fleshy axis.
In
botany a bract is a specialized leaf that is
associated with the reproductive structure
of the plant. The spadix can only
produce flowers at sexual anthesis.
True flowers contain
near microscopic sexual parts including
anthers, stamens, and stigmas. Some
aroids have tepals but others do not.
 organs mixed together
throughout the length of the spadix. To help prevent
self
pollination the female flowers are receptive prior to the
production of pollen by the male flowers. The second or unisexual
group includes those species which produce an inflorescence with
imperfect flowers which are either male or female in separated
zones.
organs mixed together
throughout the length of the spadix. To help prevent
self
pollination the female flowers are receptive prior to the
production of pollen by the male flowers. The second or unisexual
group includes those species which produce an inflorescence with
imperfect flowers which are either male or female in separated
zones. In nature an inflorescence signals that it is about to open and begins to emit an odor known scientifically as a pheromone that is attractive to selected insect pollinators. The pheromone is often released in waves so the intensity will rise and fall depending on the sexual activity of the inflorescence. When the flowers are fully receptive the pheromone (perfume) is often more intense but then wanes and rises as the chemical is released. Recent viewers of Amorphophallus titanum (Lois) at the Houston Museum of Natural Science were often curious why the scent was not always fully intense. As the event reaches the conclusion the pheromone level often drops dramatically. A photo of Lois is below.
 detect the pheromones at distances
of approximately 200 meters or over 600 feet away. The beetles
fly a zig-zag
pattern through the forest in an effort to follow the "perfume" to
its source. It is suspected there are
approximately 900 species of Cyclocephala beetles but only about one
half have been scientifically identified.
detect the pheromones at distances
of approximately 200 meters or over 600 feet away. The beetles
fly a zig-zag
pattern through the forest in an effort to follow the "perfume" to
its source. It is suspected there are
approximately 900 species of Cyclocephala beetles but only about one
half have been scientifically identified. 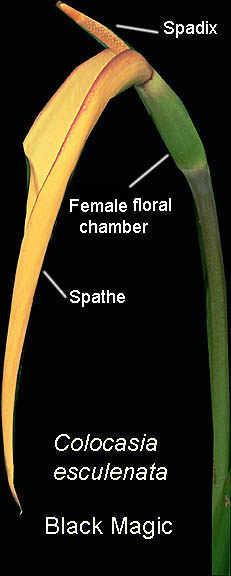 One
or several of these insect species have
evolved to be the specific or "assigned"
pollinator of any particular plant species.
Once the bodies of those "assigned"
pollinators come in contact with the
resin (if present) on the spadix (see photo,
top of page) they
collect and hold the dehisced pollen grains
from one inflorescence which is shedding
pollen at male anthesis and transport it to
another bloom at female anthesis in order to
cause the pollen to adhere to the tiny
sticky female flowers, As a result
pollination is achieved. The term dehisced
indicates pollen that has been shed.
One
or several of these insect species have
evolved to be the specific or "assigned"
pollinator of any particular plant species.
Once the bodies of those "assigned"
pollinators come in contact with the
resin (if present) on the spadix (see photo,
top of page) they
collect and hold the dehisced pollen grains
from one inflorescence which is shedding
pollen at male anthesis and transport it to
another bloom at female anthesis in order to
cause the pollen to adhere to the tiny
sticky female flowers, As a result
pollination is achieved. The term dehisced
indicates pollen that has been shed.Since many birds, bats and small animals including monkeys rely on the berries of aroids as a food source during the reproductive season the destruction of any rain forest region may adversely affect the life cycle of countless plant and animal species. Some have now either gone completely extinct or are unusually rare as the result of nothing more than the destruction of the habitat of a specific natural insect pollinator. It is also thought the natural pollinator of species such as Philodendron spiritus-sancti may have become extinct due to to forest devastation. That southeastern Brazilian Philodendron species is now incredibly rare with only six specimens known to exist in nature as a result of deforestation.
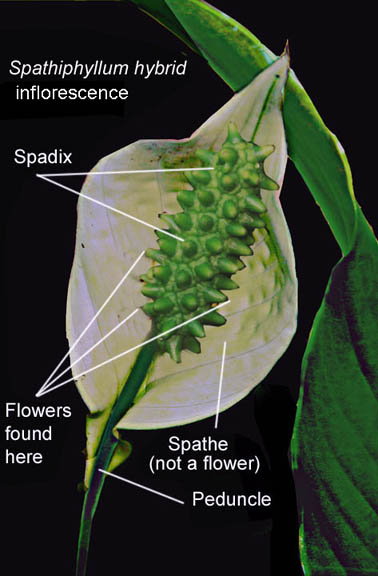
The inflorescences of Bisexual Species

 Both bisexual and unisexual aroid plant
species produce blooms consisting of a simple leaf-like and
sometimes attractively colored spathe that is attached to the base
of its attending spadix. The spadix is a spike or rod-like
structure along the length of which are hundreds of minute but
actual flowers.
Both bisexual and unisexual aroid plant
species produce blooms consisting of a simple leaf-like and
sometimes attractively colored spathe that is attached to the base
of its attending spadix. The spadix is a spike or rod-like
structure along the length of which are hundreds of minute but
actual flowers.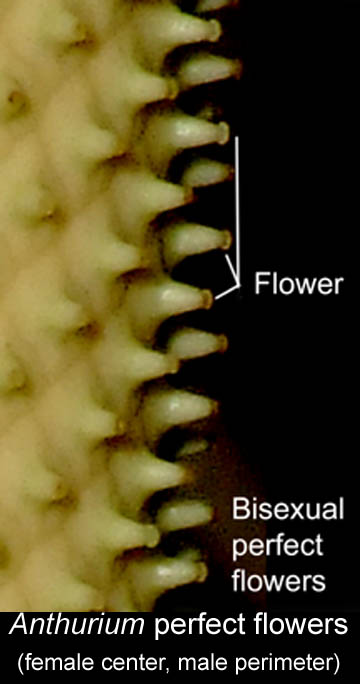 To prevent self-pollination in nature two events occur,
one following the other. The first is the initiation of female
anthesis. At this reproductive stage the female flower's stigmas
are ready to be pollinated by the pollen from another
more mature
inflorescence located on a near-by plant. That second inflorescence
would necessarily already have
reached male anthesis. After the
spathe is fully opened, a process that may take a considerable
amount of time including days or even weeks, the stigmas can be
observed to produce a tiny drop of liquid on their tips. This
liquid serves to hold the pollen grains in place once presented by
an assigned pollinator. This liquid is produced in both bisexual
and unisexual reproduction. However, within Anthurium
species it is at times possible for self pollination to occur since
some flowers begin the production of pollen before all the female
flowers are spent.
To prevent self-pollination in nature two events occur,
one following the other. The first is the initiation of female
anthesis. At this reproductive stage the female flower's stigmas
are ready to be pollinated by the pollen from another
more mature
inflorescence located on a near-by plant. That second inflorescence
would necessarily already have
reached male anthesis. After the
spathe is fully opened, a process that may take a considerable
amount of time including days or even weeks, the stigmas can be
observed to produce a tiny drop of liquid on their tips. This
liquid serves to hold the pollen grains in place once presented by
an assigned pollinator. This liquid is produced in both bisexual
and unisexual reproduction. However, within Anthurium
species it is at times possible for self pollination to occur since
some flowers begin the production of pollen before all the female
flowers are spent.An odor which is sometimes attractive and at others hardly noticeable to the human nose (but always attractive to the specific pollinator) is produced and released at this time through a process known as thermogenesis (thermo= heat, genesis= birth). That pheromone is distributed by an increase in temperature within the spadix as a result of the release of salicylic acid and other natural occurring heating of the spadix. Salicylic acid is the same chemical compound used in the production of aspirin. After female anthesis (AN-th-sis) is complete, usually in a matter of days, the stigmas dry and turn brown. u
 Shortly after this event occurs male anthesis
initiates and pollen begins to be produced.
In some species the
pollen is observed as a powdery dust that is visibly shed while in
others it appears like a fuzzy material on the spadix. Pollen
is also produced in a similar fashion within unisexual aroid
genera.
Shortly after this event occurs male anthesis
initiates and pollen begins to be produced.
In some species the
pollen is observed as a powdery dust that is visibly shed while in
others it appears like a fuzzy material on the spadix. Pollen
is also produced in a similar fashion within unisexual aroid
genera. 
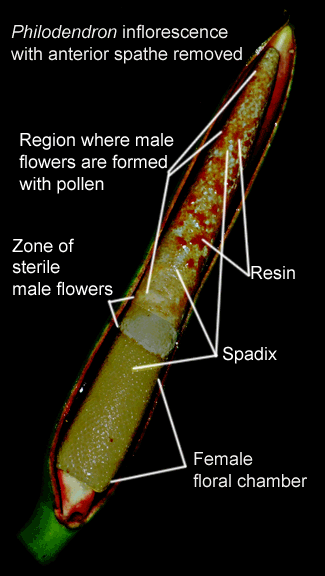
The inflorescences of Unisexual Species

 which the spathe consists of two sections,
both normally wrapped around the spadix with a constriction
separating the two sections. The upper portion of the spathe is
called the limb or blade while the lower is a convolute tube or
chamber. On the spadix the imperfect male and female flowers occur in
separate regions or zones. Normally the zone of female flowers
occurs at the very bottom of the spadix within the lowest portion of
the spathe known as the spathe tube or floral chamber. Above this female
zone at the constriction is a zone of sterile male flowers. These
sterile flowers produce a pheromone (odor) which attracts insect
pollinators in exactly the same fashion as bisexual species.
Normally, above these sterile flowers and within the upper spathes
blade or limb, occurs the zone of fertile male flowers.
The female floral chamber of some species is well hidden and often
constricted.
which the spathe consists of two sections,
both normally wrapped around the spadix with a constriction
separating the two sections. The upper portion of the spathe is
called the limb or blade while the lower is a convolute tube or
chamber. On the spadix the imperfect male and female flowers occur in
separate regions or zones. Normally the zone of female flowers
occurs at the very bottom of the spadix within the lowest portion of
the spathe known as the spathe tube or floral chamber. Above this female
zone at the constriction is a zone of sterile male flowers. These
sterile flowers produce a pheromone (odor) which attracts insect
pollinators in exactly the same fashion as bisexual species.
Normally, above these sterile flowers and within the upper spathes
blade or limb, occurs the zone of fertile male flowers.
The female floral chamber of some species is well hidden and often
constricted. The floral chamber of Alocasia species can be tightly constricted just beneath the spathe. The spathe of these species is divided into a convoluted thicker lower region which hides the female flowers. The spathe is observed to have a constriction with the female zone being roundish to globe shaped (globose to ovoid). When ready to be pollinated the restriction loosens to provide the pollinating insects access to the flower pistils. Once pollinated the zone remains on the peduncle and opens to become the fruit producing region. Once fruit begins to develop the inflorescence is then known as an infructescence (see list of definitions at the bottom of this page). Allocation are pollinated primarily by beetles and flies.
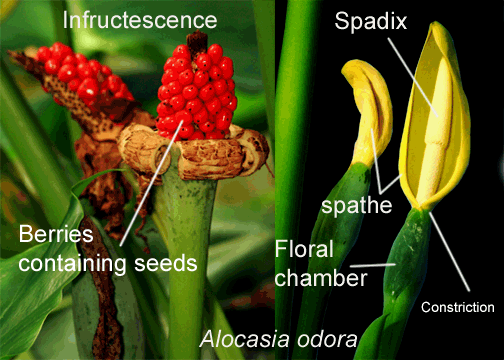
Since the majority of aroids
require a very specific insect species to do the work of
 pollination
if
that Cyclocephala beetle is not present it is unlikely the
plant will be naturally pollinated unless the species is capable of
self pollination. There is another genus
of smaller
beetles known as Neelia that visit some aroid
species but these
beetles do not
appear to feed nor mate on the inflorescence. It
appears Cyclocephala beetles
do almost all the work of
aroid pollination.
pollination
if
that Cyclocephala beetle is not present it is unlikely the
plant will be naturally pollinated unless the species is capable of
self pollination. There is another genus
of smaller
beetles known as Neelia that visit some aroid
species but these
beetles do not
appear to feed nor mate on the inflorescence. It
appears Cyclocephala beetles
do almost all the work of
aroid pollination.
The Cyclocephala pollinators are members of subfamily
Dynastinae in the family Scarabaeidae (thus
"scarab beetles) and most
belong to the genus Cyclocephala.
Some of these beetles are not particularly species specific and
visit more than a single
species or genus however
it is surmised the height of the plant from the ground may be a particular
attractant to individual beetles thus causing them not to cross
pollinate other than their naturally specified
plant species. However, human induced cross
pollination is not only possible but is frequently accomplished.
The beetles are generally drawn to the inflorescence in the late day or at dusk and are attracted by a combination of pheromones (scent) and a source of food and shelter which is composed at least in part of an oil produced on the staminate flowers containing lipids along with the enclosure of the spathe. Shelter may play a part since the male often brings along his mate in to breed at the same time. Some species have sweet smelling pheromones while others show no noticeable aroma. This aroma is produced by the sterile male flowers on the inflorescence which are attempting to entice the pollinator and to the male of that insect species the scent may be similar to the same pheromone that attracts him to a mate when she is ready to be impregnated. This point is not factually certain.

So why do they spend the night? Thermogenesis! Quite simply,
the spadix can warm enough to be noticeable to the
touch and for
the insects that may be tired from traveling long distances
 to
perform their required tasks this additional source of heat in
the rain forest creates a microclimate and may actually increase
their metabolism and encourage them to explore all
portions of
the spathe and spadix. A microclimatic zone of warmth is now
being generated within the spathe that offers both comfort and
protection along with food. This feature alone may increase the
chance of self pollination within the specimen, but another may
inhibit the same.
to
perform their required tasks this additional source of heat in
the rain forest creates a microclimate and may actually increase
their metabolism and encourage them to explore all
portions of
the spathe and spadix. A microclimatic zone of warmth is now
being generated within the spathe that offers both comfort and
protection along with food. This feature alone may increase the
chance of self pollination within the specimen, but another may
inhibit the same.
 botanists know as thermogenesis.
The presence of beetles appears to increase the temperature
produced by the event and the temperature increase appears to
increase the amount of pheromone (perfume) being exuded by
the tiny flowers. Up to 200 beetles at a single time have been
observed on a single Philodendron inflorescence during anthesis,
however,
the normal number is closer to 5 to 10. Researchers have noted
the highest temperatures appear to occur during the period when
the highest number of beetles are present. However the exact
role of thermogenesis is still not well understood.
The
pheromones (scent or perfume) produced by Philodendron
species are not always detectable to the human nose. Some
species have noticeably sweet scents in the early evening while
some exude no noticeable smell on the first day of sexual
anthesis. On other species the pheromone is noticeable
only during specific hours of the day, normally in the evening.
Most Philodendron produce their own unique pheromone
which is attractive to only a single species of pollinating
beetle.
botanists know as thermogenesis.
The presence of beetles appears to increase the temperature
produced by the event and the temperature increase appears to
increase the amount of pheromone (perfume) being exuded by
the tiny flowers. Up to 200 beetles at a single time have been
observed on a single Philodendron inflorescence during anthesis,
however,
the normal number is closer to 5 to 10. Researchers have noted
the highest temperatures appear to occur during the period when
the highest number of beetles are present. However the exact
role of thermogenesis is still not well understood.
The
pheromones (scent or perfume) produced by Philodendron
species are not always detectable to the human nose. Some
species have noticeably sweet scents in the early evening while
some exude no noticeable smell on the first day of sexual
anthesis. On other species the pheromone is noticeable
only during specific hours of the day, normally in the evening.
Most Philodendron produce their own unique pheromone
which is attractive to only a single species of pollinating
beetle.A more recent train of thought includes the likelihood infrared heat acts as an increaser to the production and distribution of the pheromones. Aroid pollination expert Dr. Marc Gibernau (GHEE-ber-no) of the University Paul Sabatier in Toulouse, France created and provided the chart below. His chart (below) shows just how "bright" the "glow" of infrared heat is the the pollinating beetle as well as indicating the increase in temperature of the spadix above the ambient temperature of the rain forest. Marc is shown standing next to the glowing inflorescence. A discussion of infrared heat and its relationship to pollination follows the chart.
 Marc suspects the
beetles are also attracted to the spadix in the darkness of the
forest due to the infrared heat produced during sexual anthesis.
In both private and public discussions with Marc in Miami, FL in
September, 2008 he explained in a presentation to the International
Aroid Society as well as to several of those of us (Julius Boos,
Christopher Rogers and Steve Lucas) individually there is a significant increase
in temperature above
the ambient temperature of the rain forest at night once the
inflorescence reaches anthesis. The average Philodendron temperature increase is approximately 12 degrees
Celsius (a 21.6 degree Fahrenheit increase) above ambient but a few plant species
can increase in temperature by as much as 20 degrees C (36 degrees F)
above ambient.
The heat can be so intense it can be felt on the palm of an opened hand held in front
of the spadix which we have personally experienced. On the chart
below the
spadix shown is
Philodendron solimoesense. If you notice the temperature
gradients you will see the spadix of Philodendron solimoesense
increases in temperature 14 degrees C or 25.2 degrees F above
the surrounding rain forest ambient temperature.
Marc suspects the
beetles are also attracted to the spadix in the darkness of the
forest due to the infrared heat produced during sexual anthesis.
In both private and public discussions with Marc in Miami, FL in
September, 2008 he explained in a presentation to the International
Aroid Society as well as to several of those of us (Julius Boos,
Christopher Rogers and Steve Lucas) individually there is a significant increase
in temperature above
the ambient temperature of the rain forest at night once the
inflorescence reaches anthesis. The average Philodendron temperature increase is approximately 12 degrees
Celsius (a 21.6 degree Fahrenheit increase) above ambient but a few plant species
can increase in temperature by as much as 20 degrees C (36 degrees F)
above ambient.
The heat can be so intense it can be felt on the palm of an opened hand held in front
of the spadix which we have personally experienced. On the chart
below the
spadix shown is
Philodendron solimoesense. If you notice the temperature
gradients you will see the spadix of Philodendron solimoesense
increases in temperature 14 degrees C or 25.2 degrees F above
the surrounding rain forest ambient temperature.
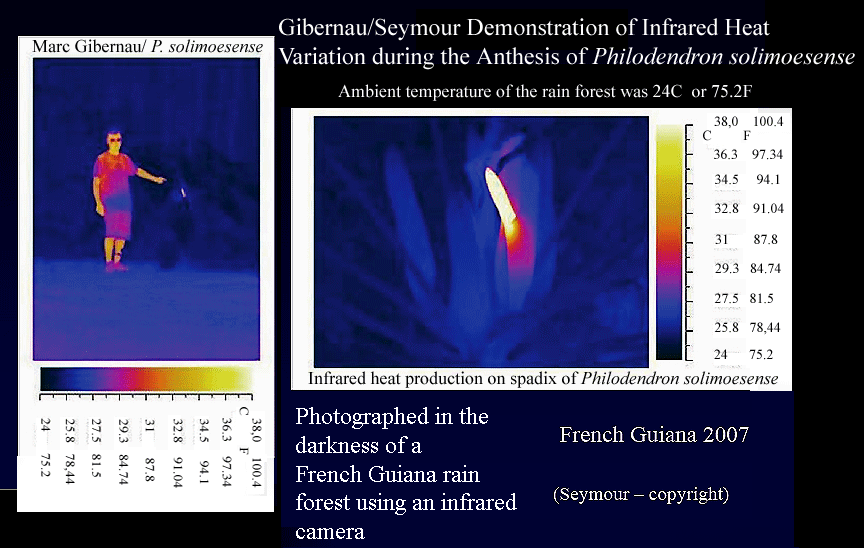
 In photographic documentation
seen on the chart (above) created with an infrared camera the "glow" of a sexually mature
Philodendron solimoesense
spadix is "visible" and Marc theorizes the beetles can detect that
infrared heat with a method similar to a pilot seeing the glow of a
runway light at night. Since the beetle uses the spathe and
spadix as a source of food (pollen) and a place for warmth during
its own sexual
reproduction the "glow" is an open invitation to fly
to that source of food, shelter and warmth.
In photographic documentation
seen on the chart (above) created with an infrared camera the "glow" of a sexually mature
Philodendron solimoesense
spadix is "visible" and Marc theorizes the beetles can detect that
infrared heat with a method similar to a pilot seeing the glow of a
runway light at night. Since the beetle uses the spathe and
spadix as a source of food (pollen) and a place for warmth during
its own sexual
reproduction the "glow" is an open invitation to fly
to that source of food, shelter and warmth.
At present, Marc and his associates are working to prove the beetle does not actually "see" the infrared heat, but instead detects it with receptors on their antennae or bodies instead of seeing it with their eyes.
Marc forwarded these additional comments in a
personal email received on October 14, 2008:
"My picture (below) is an inflorescence of Philodendron solimoesense.
Your
paragraph about anthesis may also need to explain the time
shift between male and female phases (protogyny) because people may
think
an inflorescence can self-pollinate. On day one the
female flowers are receptive and the next day the male flowers
produce pollen because the flower are
synchronized. On the first day the
inflorescence is at the female stage and all stigma are
receptive
 for fecundation/pollination by pollen grains. On the next
day (2nd day), the stigma are no longer receptive but the anthers
are fully ripe (mature) and shed the pollen. In a few species,
such as
in some Anthurium, these two sexual phases are overlapping and
self-pollination may occur. Hence, aroids need pollen vectors
(insects) for pollination between different inflorescences.
The Cyclocephala beetles carry
the pollen from a male-stage inflorescence to a female-stage inflorescence.
From the standpoint of a botanist the aroid reproductive structure would be the
same as to view
the inflorescence, which is composed of many flowers packed together,
to functionally
"behaves" like a flower."
As a result of the
Cyclocephala beetle bringing pollen from one aroid
species to
another aroid of the same species the specimen is able to grow viable seeds.
for fecundation/pollination by pollen grains. On the next
day (2nd day), the stigma are no longer receptive but the anthers
are fully ripe (mature) and shed the pollen. In a few species,
such as
in some Anthurium, these two sexual phases are overlapping and
self-pollination may occur. Hence, aroids need pollen vectors
(insects) for pollination between different inflorescences.
The Cyclocephala beetles carry
the pollen from a male-stage inflorescence to a female-stage inflorescence.
From the standpoint of a botanist the aroid reproductive structure would be the
same as to view
the inflorescence, which is composed of many flowers packed together,
to functionally
"behaves" like a flower."
As a result of the
Cyclocephala beetle bringing pollen from one aroid
species to
another aroid of the same species the specimen is able to grow viable seeds.
I asked Marc how the beetles find the inflorescence in the dark of the forest and was told the two attractants appear to work together. First, since the pheromone produced by inflorescence can travel on the wind for 200 meters or so the beetles apparently first detect the scent in the wind. But since the wind shifts through the forest they have to fly a zig zag pattern back and forth in to follow it to the source. Once they are close enough to "see" the "glow" of the infrared heat they are drawn to the source in the same way a pilot sees his destination runway and simply follows the "lights" home.
You can find more information on Philodendron species by clicking this link.
Forced commercial inflorescence inducement
Aroids
produce their inflorescences depending on the time of year and season
nature has predetermined. Some species produce a spathe and spadix only during
the dry or wet season in their natural habitat while others freely
produce an inflorescence any time of the year. Hybridized
forms of Anthurium andreanum (Flamingo Flower) as well as hybridized
Spathiphyllum (Peace Lilies) commonly sold in garden centers and
discount centers can be
seen with an
 open inflorescence virtually any month of the year.
open inflorescence virtually any month of the year.
The commercial growers that produce these plants use a chemical known as
gibberellic acid often sold as GA3 to induce the plants to produce an inflorescences in
order to make them more saleable at the time a buyer sees one in the
store. Gibberellic acid is natural plant hormone and is used
in agriculture to stimulate both cell division and cell elongation that
affects the leaves as well as stems of a plant. The continued use
of the chemical in agriculture eventually affects fruit development. Since the
fruit of an aroid is produced on the spadix gibberellic acid thus speeds
up the the production
of inflorescences.
Gibberellic
acid is used commercially to make all the plants in one group bloom
at the same time. Through repeated use these large growers force
the natural growth of a spathe and spadix regardless of season.
They have
calculated the quantity to be used and know how much
gibberellic acid to
apply to any particular
species but these formulations are often guarded secrets.
An article by Dr. Paul Resslar on the production of an inflorescence of Caladium humboldtii can be found in the IAS journal Aroideana, volume 31, where he discusses the amounts and methods of application used in his research. In Dr. Resslar's experience the chemical also can result in deformities. Deformity is considered a minor problem in commercial aroid production including double spathes, spadices with strange shapes, malformed leaves and other side effects. One of the side effects may well be the plant becomes dependent on the chemical to induce the production of an inflorescence and flowers. A presentation by aroid enthusiast Ted Held at the 2008 International Aroid Show in Miami pointed out these exact problems.
Discussions on these effects can be found on the International Aroid
Society forum Aroid l by searching the archives of the forum on the
internet. Some of the world's best aroid botanists and experts do
not find the use of the chemical to be wise. As a result, the use of
gibberellic acid by aroid collectors
with no experience in chemical use is not
advisable and
 should be used with caution even though it will work. The use is primarily not
advised since collectors have no way of knowing how
much of the chemical to apply to any particular specimen based on species
or size. But there are other problems. The instructions on some of
the containers of GA3 say to apply the chemical near the base of the
plant but it also says boldly not to allow the chemical to come in
contact with the roots! Some material indicates this may not be
accurate but the warning can be found on sometimes on the product
itself. Since the roots are at the base of the
plant is is obvious the chemical can be dangerous to your plants.
Other potential problems
stated in papers on the commercial use of GA3 indicate the chemical will
increase plant height, slightly reduce leaf width, and soften stems
during periods of low light during shorter days. Still, commercial
growers love the product because it increases their production and
profit.
should be used with caution even though it will work. The use is primarily not
advised since collectors have no way of knowing how
much of the chemical to apply to any particular specimen based on species
or size. But there are other problems. The instructions on some of
the containers of GA3 say to apply the chemical near the base of the
plant but it also says boldly not to allow the chemical to come in
contact with the roots! Some material indicates this may not be
accurate but the warning can be found on sometimes on the product
itself. Since the roots are at the base of the
plant is is obvious the chemical can be dangerous to your plants.
Other potential problems
stated in papers on the commercial use of GA3 indicate the chemical will
increase plant height, slightly reduce leaf width, and soften stems
during periods of low light during shorter days. Still, commercial
growers love the product because it increases their production and
profit.
The next time you buy a beautiful Anthurium or Spathiphyllum at a discount store and find it begins to produce odd shaped leaves and spathes or rarely if ever produces an inflorescence once you get it home there may be a reason. The specimen has very likely been fed gibberellic acid since it was nearing sexual maturity to force it to mature and bloom early. Without the constant use of the hormone to induce inflorescence production the specimen cannot get its "fix" and as a result may rarely bloom again. It has been "hooked" on the chemical! Collectors should consider using the product with extreme caution.
Aroideana is the journal of the International Aroid Society and out of print copies can be ordered at http://www.aroid.org/
Artificial pollination
 To achieve hand-pollination the pollen from
a bloom of the same or a related species which is already at
male anthesis must be transported. In nature the event is completed
by an insect pollinator that has been attracted by the bloom's
odor but within cultivation it must be done manually. The hand of
man must perform the tasks of collecting the pollen and transferring
it
from one inflorescence which is experiencing male anthesis to
another younger inflorescence which is at female
anthesis.
To achieve hand-pollination the pollen from
a bloom of the same or a related species which is already at
male anthesis must be transported. In nature the event is completed
by an insect pollinator that has been attracted by the bloom's
odor but within cultivation it must be done manually. The hand of
man must perform the tasks of collecting the pollen and transferring
it
from one inflorescence which is experiencing male anthesis to
another younger inflorescence which is at female
anthesis.  plant which is now undergoing female
anthesis. If the pollination is successful the inflorescence will
"hold" on the second plant and the fruit will begin to develop. If the
artificial pollination is truly a success this will become obvious
as the female flowers enlarge to produce berries. Each berry
ranging in color from red to orange through yellow, purple or white
may contain a single large seed or several medium sized seeds which
are surrounded by a pulp and skin which is often tasty depending on
the genus and species involved. The
development time may be long and in some
species may take more than a
year from pollination to ripeness and seed maturity.
plant which is now undergoing female
anthesis. If the pollination is successful the inflorescence will
"hold" on the second plant and the fruit will begin to develop. If the
artificial pollination is truly a success this will become obvious
as the female flowers enlarge to produce berries. Each berry
ranging in color from red to orange through yellow, purple or white
may contain a single large seed or several medium sized seeds which
are surrounded by a pulp and skin which is often tasty depending on
the genus and species involved. The
development time may be long and in some
species may take more than a
year from pollination to ripeness and seed maturity.  Hand-pollination of any aroid species can be
accomplished relatively simply by collecting pollen from one bloom
using a small brush such as a camel hair artist brush wetted with
sterile water and transferring this pollen to a bloom as soon as it
is observed the second is beginning to open. Be careful to observe
for the sticky liquid exuded by the female flowers once ready to
accept the pollen. Coating the female flowers with as much pollen
as possible will act to ensure the pollen "holds" and the
reproductive cycle is complete.
Hand-pollination of any aroid species can be
accomplished relatively simply by collecting pollen from one bloom
using a small brush such as a camel hair artist brush wetted with
sterile water and transferring this pollen to a bloom as soon as it
is observed the second is beginning to open. Be careful to observe
for the sticky liquid exuded by the female flowers once ready to
accept the pollen. Coating the female flowers with as much pollen
as possible will act to ensure the pollen "holds" and the
reproductive cycle is complete. 

For Information on joining the International Aroid Society please click this link.
Want to see the development of a Philodendron spathe and spadix?
Watch pollen develop! 78 Days of the
Anthurium regale
reproductive cycle.
http://www.exoticrainforest.com/Anthurium%20regale%20pc.html
|
Terms you may encounter in further research:
Anther - (AN-ther)
The part of the stamen where the pollen is produced and contained
Anthesis
- (an-THE-sis)
The period during which a flower is fully open and functional. The time of pollination or bloom.
Inflorescence - (in-flor-ES-ence)
A group or cluster of flowers on a branch of a plant, in botany used to describe the structure of a spathe and spadix
Infructescence - (in-fruc-TES-sence)
Nectar
- (NEC-tar)
Perigone - (PERU-gown)
Pheromones
(fer-UH-mones)
A scent produced by the spadix during anthesis to attract insect pollinators and distribute pollen. Pistil - (PIS-til)
The female, ovule-bearing organ of a flower,
including the stigma, style, and ovary
Pollen - (POL-in)
sometimes incorrectly called flower sperm, is a fine to coarse powder consisting of pollen grains which produce the male gametes (sperm cells) of seed plants
Stamen - (STA-men)
Male reproductive parts of a flower
Stigma - (STIG-ma)
Stigmatic Hairs
(stig-MAT-ic)
|